Жизнь молекул днк. Днк и гены Свойства молекулы днк кратко
Жизнь ДНК (дезоксирибонуклеиновых кислот)
Определение понятия "ДНК"
Ген - это совокупность сегментов ДНК, обуславливающих образование либо молекулы РНК, либо белкового продукта (Сингер М., Берг П., 1998).
У человека около 30000 генов. Во всём объёме ДНК структурные гены (т.е. те, которые кодируют белки, идущие на построение стуктур организма) занимают лишь 3-10%.
Наименьшая функциональная единица ДНК состоит из следующих элементов: структурный ген, регуляторные зоны, регуляторные гены.
Строение молекулы ДНК
Молекулы ДНК имеют вид длинных двойных цепей полимеров – полинуклеотидов, состоящих из мономеров – нуклеотидов. Двойная цепь закручена в спираль. Поэтому ДНК похожа на винтовую лестницу (посмотрите на рисунок вверху). Каждый нуклеотид включает одно из четырех азотистых оснований – аденин (А), гуанин (Г), цитозин (Ц) или тимин (Т), одну молекулу пентозы (пятиуглеродный сахар) и один остаток фосфорной кислоты. Обычно молекула ДНК состоит из двух комплементарных нитей, которые образуют двойную спираль. При этом аденин одной нити находится в паре с тимином другой (стабилизируется двумя водородными связями), а гуанин аналогично связан с цитозином (тремя водородными связями). Последовательность азотистых оснований в молекуле ДНК несет информацию, необходимую для синтеза белков. ДНК - очень длинные молекулы, состоящие из множества нуклеотидов. Например, геном человека состоит из 46 хромосом, основу которых составляют молекулы ДНК, которые в совокупности собраны примерно из 3 млрд нуклеотидны пар.
У эукариот генетический материал находится в ядре клетки в хромосомах. Хромосомы в активном состоянии существуют в виде хроматина. Хроматин содержит около 40% ДНК, 40% гистонов (щелочных белков), около 20% негистоновых хромосомных белков и немного РНК.
Видео: Строение хромосомы
ДНК мы можем отнести к "живым системам", к "живым молекулам" на том основании, что они лежат в основе жизни вообще, а также обладают рядом важнейших свойств живого, в частности, способностью к размножению. ДНК насктолько самостоятельны и самодостаточны, что способны существовать даже вне клетки - в виде вирусов. В своей жизни молекулы ДНК проходят жизненные этапы, напоминающие нам жизнь более сложных биологических систем - живых организмов. Это такие этапы как рождение, созревание, работа (деятельность) и "смерть".
Тема: Строение ДНК
Домашнее задание
- Знать и уметь писать структурные формулы нуклеотидов: А, Т, Г, Ц, У.
- Знать устройство молекул ДНК и их организацию в хромосомы.
- Знать способы связывания нуклеотидов в ДНК по вертикали и горизонтали. Понятие о 3"-5" связях.
- Уметь пользоваться таблицей генетического кода для построения молекул пептидов на основе участка ДНК размером от 12 и более нуклеотидов.
Видео: Хромосомы, митоз, репликация
Этапы жизни молекулы ДНК
Рождение (репликация) - созревание (хромосомы) - работа (транскрипция) - управление (регуляция) - видоизменение (мутация) - "смерть"
1. Репликация ДНК - рождение новой дочерней нити ДНК на родительской нити.
2. Созревание ДНК - формирование хромосомы.
3. Транскрипция ДНК - работа ДНК в виде матричного синтеза на ней РНК.
4. Регуляция транскрипции - управление деятельностью ДНК по транскрипции.
5. Репарация ДНК - восстановление повреждённых участков.
6. Изменения структуры ДНК - мутации, транспозоны.
7. Деградация ДНК - разрушение при каждом цикле репликации.
1. Рождение - репликация
Репликация ДНК проходит очень просто, на счёт "раз, два, три", то есть в три этапа: 1) инициация, 2) элонгация, 3) терминация.
1. Инициация - начинание
Мишень для запуска репликации
Репликация огромной молекулы ДНК начинается с возникновения репликативной точки. Эта точка имеет специфическую последовательность богатую парами А-Т. Такие учкастки в ДНК как раз и являются мишенями для белков, инициирующих репликацию. Именно к ним присоединяются специальные распознающие белки, которые обеспечивают присоединение ферментов репликации хеликазы
и топоизомеразы
(гиразы) и таким образом запускают процесс репликации. Хеликаза
расплетает ДНК на две цепи. Образуется репликативная вилка. Молекула ДНК жестко закреплена на ядерном матриксе и не может свободно вращаться при расплетании какого-либо участка. Это блокирует продвижение хеликазы по цепи. Топоизомераза надрезает нити ДНК и снимает структурное напряжение.
В одной репликативной вилке действуют две хеликазы, которые движутся в противоположных направлениях. Разделенные цепи фиксируются ДНК- связывающими белками. Участки формирования репликативной вилки называются «точками ori» (origin - начало). У эукариот одновременно образуется тысячи таких вилок, что обеспечивает высокую скорость репликации.
2. Элонгация - продолжение (удлиннение)
Наращивание дочерних цепей ДНК на двух родительских цепях происходит неодинаково. ДНК- полимераза III прокариот и δ- или α-ДНК-полимеразы эукариот могут осуществлять синтез новой цепи ДНК лишь в направлении 5’>3’, т.к. могут присоединить новый нуклеотид только к углероду в положении 3’, но не в положении 5’.
Цепь с такой направленностью называется лидирующей . На ней синтез дочерней нити ДНК идёт непрерывно. ДНК-полимераза III или δ-полимераза непрерывно присоединяют к ней комплементарные нуклеотиды.
Цепь с полярностью 3’>5’ является отстающей и достраивается по частям (также в направлении 5’>3’). α-ДНК-полимераза (или ДНК-полимераза III) синтезирует на этой цепи короткие участки - фрагменты Оказаки.
Синтез фрагментов Оказаки и лидирующей цепи начинается с образования РНК-праймеров (затравок ) длиной 10-15 рибонуклеотидов ферментом праймазой (РНК-полимеразой). Ни одна из ДНК-полимераз не способна начать синтез ДНК с нуля, а может лишь достраивать существующую цепь. Параллельно с образованием лидирующей цепи или фрагментов Оказаки происходит удаление рибонуклеотидов из праймеров и замена их нуклеотидами ДНК. Замена рибонуклеиновых участков (праймеров) на участки ДНК происходит с помощью β-ДНК-полимеразы, которая имеет как экзонуклеазную, так и полимеразную активность.
Таким образом, репликация невозможна без частичной временной транскрипции.
Скорость репликации (элонгации) ДНК составляет около 45000 нуклеотидов в минуту, таким образом, родительская вилка расплетается со скоростью 4500 об/мин. Это сопоставимо, например, со скоростью вращения охлаждающего вентилятора в компьютере (1300-4800 об/мин).
3. Терминация - завершение (окончание)
Завершение репликации происходит тогда, когда пробелы между фрагментами Оказаки заполнятся нуклеотидами (при участии ДНК-лигазы) с образованием двух непрерывных двойных цепей ДНК и когда встретятся две репликативные вилки. Затем происходит закручивание синтезированных ДНК с образованием суперспиралей.
Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и действием ДНК-полимераз, которые обладают кроме полимеразной, еще и экзонуклеазной активностью и способны распознавать и исправлять ошибки. Если включается некомплементарный нуклеотид, то фермент делает шаг назад, отщепляет его и продолжает полимеразную реакцию. Поэтому процесс репликации является высокоточным.
После завершения репликации происходит метилирование ДНК в участках –GАТС- по аденину (с образованием N-метиладенина) и остаткам цитозина с образованием 5-метилцитозина. Метилирование не нарушает комплементарности цепей и является необходимым для формирования структуры хромосом и регуляции транскрипции генов.
У прокариот, таких как бактерии, ДНК способна реплицироваться, не распрямляясь в линейную молекулу, то есть оставаясь в характерной для неё кольцевой форме.
Видео: П
2. Созревание - формирование хромосомы и хроматина
3. Работа - транскрипция
Видео: Блокировка работы гена
4. Управление - регуляция
5. Восстановление (починка) - репарация
6. Видоизменение - мутация .
7. "Смерть" - деградация при репликации.
Мономерными звеньями которого являются нуклиатиды.
Что такое ДНК?
Вся информация о строении и функционировании любого живого организма содержится в закодированном виде в его генетическом материале. Основу генетического материала организма составляет дезоксирибонуклеиновая кислота (ДНК) .
ДНК большинства организмов – это длинная двухцепочечная полимерная молекула. Последовательность мономерных звеньев (дезоксирибонуклеотидов ) в одной ее цепи соответствует (комплементарна ) последовательности дезоксирибонуклеотидов в другой. Принцип комплементарности обеспечивает синтез новых молекул ДНК, идентичных исходным, при их удвоении (репликации ).
Участок молекулы ДНК, кодирующий определенный признак, – ген .
Гены – это индивидуальные генетические элементы, имеющие строго специфичную нуклеотидную последовательность, и кодирующие определенные признаки организма. Одни из них кодируют белки, другие - только молекулы РНК.
Информация, которая содержится в генах, кодирующих белки (структурных генах), расшифровывается в ходе двух последовательных процессов:
- синтеза РНК (транскрипции ): на определенном участке ДНК как на матрице синтезируется матричная РНК (мРНК).
- синтеза белка (трансляции): В ходе согласованной работы многокомпонентной системы при участии транспортных РНК (тРНК ), мРНК , ферментов и различных белковых факторов осуществляется синтез белковой молекулы .
Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы определяет ее структуру и функции.
Строение ДНК
ДНК – это линейный органический полимер . Его – нуклеотиды , которые, в свою очередь, состоят из:
При этом, фосфатная группа присоединена к 5′-атому углерода моносахаридного остатка, а органическое основание - к 1′-атому .
Основания в ДНК бывают двух типов:

Строение нуклеотидов в молекуле ДНК
В ДНК моносахарид представлен 2′-дезоксирибозой , содержащей только 1 гидроксильную группу (ОН) , а в РНК - рибозой , имеющей 2 гидроксильные группы (OH ).
Нуклеотиды соединены друг с другом фосфодиэфирными связями , при этом фосфатная группа 5′-углеродного атома одного нуклеотида связана с З’-ОН-группой дезоксирибозы соседнего нуклеотида (рисунок 1). На одном конце полинуклеотидной цепи находится З’-ОН-группа (З’-конец), а на другом - 5′-фосфатная группа (5′-конец).
Уровни структуры ДНК
Принято выделять 3 уровня структуры ДНК:
- первичную;
- вторичную;
- третичную.
Первичная структура ДНК – это последовательность расположения нуклеотидов в полинуклеотидной цепи ДНК.
Вторичная структура ДНК стабилизируется между комплементарными парами оснований и представляет собой двойную спираль из двух антипараллелных цепочек, закрученных вправо вокруг одной оси.
Общий виток спирали- 3,4нм , расстояние между цепочками 2нм.
Третичная структура ДНК – суперсперализация ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы, что часто вызвано ковалентным соединением их открытых концов. Суперспиральная структура ДНК обеспечивает экономную упаковку очень длинной молекулы ДНК в хромосоме. Так, в вытянутой форме длина молекулы ДНК составляет 8 см , а в форме суперспирали укладывается в 5 нм .
Правило Чаргаффа
Правило Э. Чаргаффа – это закономерность количественного содержания азотистых оснований в молекуле ДНК:
- У ДНК молярные доли пуриновых и пиримидиновых оснований равны: А+ G = C + Т или (А + G )/(C + Т)=1 .
- В ДНК количество оснований с аминогруппами (А + C ) равно количеству оснований с кетогруппами (G + Т): А + C = G + Т или (А + C )/(G + Т)= 1
- Правило эквивалентности, то есть: А=Т, Г=Ц; А/Т = 1; Г/Ц=1.
- Нуклеотидный состав ДНК у организмов различных групп специфичен и характеризуется коэффициентом специфичности: (Г+Ц)/(А+Т). У высших растений и животных коэффициент специфичности меньше 1, и колеблется незначительно: от 0,54 до 0,98 , у микроорганизмов он больше 1.
Модель ДНК Уотсона-Крика
Б 1953 г. Джеймс Уотсон и Фрэнсис Крик , основываясь на данных рентгеноструктурного анализа кристаллов ДНК, пришли к выводу, что нативная ДНК состоит из двух полимерных цепей, образующих двойную спираль (рисунок 3).
Навитые одна на другую полинуклеотидные цепи удерживаются вместе водородными связями , образующимися между комплементарными основаниями противоположных цепей (рисунок 3). При этом аденин образует пару только с тимином , а гуанин - с цитозином . Пара оснований А-Т стабилизируется двумя водородными связями , а пара G-С - тремя .
Длина двухцепочечной ДНК обычно измеряется числом пар комплементарных нуклеотидов (п .н .). Для молекул ДНК, состоящих из тысяч или миллионов пар нуклеотидов, приняты единицы т.п.н. и м.п.н. соответственно. Например, ДНК хромосомы 1 человека представляет собой одну двойную спираль длиной 263 м.п.н .
Сахарофосфатный остов молекулы , который состоит из фосфатных групп и дезоксирибозных остатков, соединенных 5’-З’-фосфодиэфирными связями , образует «боковины винтовой лестницы», а пары оснований А-Т и G-С - ее ступеньки (рисунок 3).
Рисунок 3: Модель ДНК Уотсона-Крика
Цепи молекулы ДНК антипараллельны : одна из них имеет направление 3’→5′ , другая 5’→3′ . В соответствии с принципом комплементарности , если в одной из цепей имеется нуклеотидная последовательность 5-TAGGCAT-3′ , то в комплементарной цепи в этом месте должна находиться последовательность 3′-ATCCGTA-5′ . В этом случае двухцепочечная форма будет выглядеть следующим образом:
- 5′-TAGGCAT-3′
- 3-ATCCGTA-5′.
В такой записи 5′-конец верхней цепи всегда располагают слева, а 3′-конец - справа.
Носитель генетической информации должен удовлетворять двум основным требованиям: воспроизводиться (реплицироваться) с высокой точностью и детерминировать (кодировать) синтез белковых молекул .
Модель ДНК Уотсона-Крика полностью отвечает этим требованиям, так как:
- согласно принципу комплементарности каждая цепь ДНК может служить матрицей для образования новой комплементарной цепи. Следовательно, после одного раунда образуются две дочерние молекулы, каждая из которых имеет такую же нуклеотидную последовательность, как исходная молекула ДНК.
- нуклеотидная последовательность структурного гена однозначно задает аминокислотную последовательность кодируемого ею белка.
- Одна молекула ДНК человека вмещает порядка 1,5 гигабайта информации . При этом, ДНК всех клеток человеческого организма занимают 60 млрд. терабайт, что сохраняются на 150-160 граммах ДНК.
- Международный день ДНК отмечается 25 апреля. Именно в этот день в 1953 году Джеймс Уотсон и Фрэнсис Крик опубликовали в журнале Nature свою статью под названием «Молекулярная структура нуклеиновых кислот» , где описали двойную спираль молекулы ДНК.
Список литературы: Молекулярная биотехнология: принципы и применение, Б.Глик, Дж. Пастернак, 2002 год
Химический состав ДНК и её макромолекулярная организация. Типы спиралей ДНК. Молекулярные механизмы рекомбинации, репликации и репарации ДНК. Понятие о нуклеазах и полимеразах. Репликация ДНК как условие передачи генетической информации потомкам. Общая характеристика процесса репликации. Действия, происходящие в вилке репликации. Репликация теломеров, теломераза. Значение недорепликации конечных фрагментов хромосом в механизме старения. Системы исправления ошибок репликации. Корректорские свойства ДНК-полимераз. Механизмы репарации поврежденной ДНК. Понятие о заболеваниях репарации ДНК. Молекулярные механизмы общей генетической рекомбинации. Сайт-специфическая рекомбинация. Генная конверсия.
В 1865г. Грегор Мендель открыл гены, а его современник Фридрих Мишер в 1869г. открыл нуклеиновые кислоты (в ядрах клеток гноя и сперматозоидов лосося). Однако долго еще эти открытия не связывали друг с другом, долго еще структуру и природу вещества наследственности не знали. Генетическая роль НК была установлена после открытия и объяснения явлений трансформации (1928, Ф.Гриффитс; 1944, О. Эвери), трансдукции (1951, Ледерберг, Циндер) и размножения бактериофагов (1951, А. Херши, М. Чейз).
Трансформация, трансдукция и размножение бактериофагов убедительно доказали генетическую роль ДНК. У РНК - содержащих вирусов (СПИДа, гепатита В, гриппа, ВТМ, лейкоза мышей и др.) эту роль выполняет РНК.
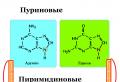
Строение нуклеиновых кислот . НК - биополимеры, участвующие в хранении и передаче генетической информации. Мономеры НК - нуклеотиды, состоящие из азотистого основания, моносахарида и одной или нескольких фосфатных групп. В составе НК все нуклеотиды являются монофосфатами. Нуклеотид без фосфатной группы называется нуклеозидом. Сахар, входящий в состав НК, представляет собой D-изомер и β-аномер рибозы или 2-дезоксирибозы. Нуклеотиды, содержащие рибозу, называются рибонуклеотидами и являются мономерами РНК, а нуклеотиды - производные дезоксирибозы, являются дезоксирибонуклеотидами, и из них состоит ДНК. Азотистые основания бывают двух типов: пурины - аденин, гуанин и пиримидины - цитозин, тимин, урацил. В состав РНК и ДНК входят аденин, гуанин, цитозин; урацил встречается только в РНК, а тимин только в ДНК.
В ряде случаев в НК присутствуют редко встречающиеся минорные нуклеотиды, такие как дигидроуридин, 4-тиоуридин, инозин и др. Разнообразие их особенно велико у тРНК. Минорные нуклеотиды образуются в результате химических превращений оснований НК, происходящих уже после образования полимерной цепи. Чрезвычайно распространены в РНК и ДНК различные метилированные производные: 5-метилуридин, 5-метилцитидин, l-N-метиладенозин, 2-И-метилгуанозин. У РНК объектом метилирования могут быть и 2"-гидроксигруппы остатков рибозы, что приводит к образованию 2"-О-метилцитидина или 2"-О-метилгуанозина.
Рибонуклеотидные и дезоксирибонуклеотидные звенья соединяются между собой с помощью фосфодиэфирных мостиков, связывающих 5"-гидроксильную группу одного нуклеотида с 3"-гидроксильной группой следующего. Таким образом, регулярная основная цепь образована фосфатными и рибозными остатками, а основания присоединены к сахарам подобно тому, как присоединены боковые группы в белках. Порядок следования оснований вдоль цепи называется первичной структурой НК. Последовательность оснований принято читать в направлении от 5"- к 3"- углеродному атому пентозы.
Структура ДНК. Модель структуры ДНК в виде двойной спирали была предложена Уотсоном и Криком в 1953 г (рис.7).
Согласно этой трехмерной модели, молекула ДНК состоит из двух противоположно направленных полинуклеотидных цепей, которые относительно одной и той же оси образуют правую спираль. Азотистые основания находятся внутри двойной спирали, и их плоскости перпендикулярны основной оси, а сахарофосфатные остатки экспонированы наружу. Между основаниями образуются специфические Н-связи: аденин - тимин (или урацил), гуанин - цитозин, получившие название уотсон-криковского спаривания. В результате более объемные пурины всегда взаимодействуют с пиримидинами, имеющими меньшие размеры, что обеспечивает оптимальную геометрию остова. Антипараллельные цепи двойной спирали не являются идентичными ни по последовательности оснований, ни по нуклеотидному составу, но они комплементарны друг другу именно благодаря наличию специфического водородного связывания между указанными выше основаниями.
Комплементарность очень важна для копирования (репликации) ДНК. Соотношения между числом различных оснований в ДНК, выявленные
Рис.7. В - форма ДНК
Чарграффом с соавт. в 50-х гг., имели большое значение для установления структуры ДНК: было показано, что число адениновых остатков в основаниях цепи ДНК, независимо от организма, равно числу тиминовых, а число гуаниновых - числу цитозиновых. Эти равенства являются следствием избирательного спаривания оснований (рис.8).
Геометрия двойной спирали такова, что соседние пары оснований находятся друг от друга на расстоянии 0.34 нм и повернуты на 36° вокруг оси спирали. Следовательно, на один виток спирали приходится 10 пар оснований, и шаг спирали равен 3.4 нм. Диаметр двойной спирали равен 20 нм и в ней образуются два желобка - большой и малый. Это связано с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем азотистые основания.
Стабильность структуры ДНК обусловлена разными типами взаимодействия, среди которых основными являются Н-связи между основаниями и межплоскостное взаимодействие (стэкинг). Благодаря последнему обеспечиваются не только выгодные ван-дер-ваальсовы контакты между атомами, но и возникает

Рис.8. Принцип комплементарности и антипараллельности цепей ДНК
дополнительная стабилизация вследствие перекрывания р-орбиталей атомов параллельно расположенных оснований. Стабилизации способствует также благоприятный гидрофобный эффект, проявляющийся в защищенности малополярных оснований от непосредственного контакта с водной средой. Напротив, сахарофосфатный остов с его полярными и ионизированными группами экспонирован, что также стабилизирует структуру.
Для ДНК известны четыре полиморфные формы: А, В, С и Z. Обычной структурой является В-ДНК, в которой плоскости пар оснований перпендикулярны оси двойной спирали (рис.7.). В А-ДНК плоскости пар оснований повернуты примерно на 20° от нормали к оси правой двойной спирали; на виток спирали здесь приходится 11 пар оснований. В С-ДНК на витке спирали 9 пар оснований. Z-ДНК - это левая спираль с 12 парами оснований на виток; плоскости оснований примерно перпендикулярны оси спирали. ДНК в клетке обычно находится в В-форме, но отдельные ее участки могут находиться в A, Z или даже в иной конформации.
Двойная спираль ДНК не застывшее образование, она находится в постоянном движении:
· деформируются связи в цепях;
· раскрываются и закрываются комплементарные пары оснований;
· ДНК взаимодействует с белками;
· если напряжение в молекуле велико, то она локально расплетается;
· правая спираль переходит в левую.
Различают 3 фракции ДНК:
1.Частоповторяемая (сателлитная) – до 106 копий генов (у мыши 10%). Она не участвует в синтезе белка; разделяет гены; обеспечивает кроссинговер; содержит транспозоны.
2.Слабоповторяемая – до 102 - 103 копий генов (у мыши 15%). Содержит гены синтеза т-РНК, гены синтеза белков рибосом и белков хроматина.
3.Уникальная (неповторяемая) – у мыши 75% (у человека 56%). Состоит из структурных генов.
Локализация ДНК: 95 % ДНК локализуется в ядре в хромосомах (линейные ДНК) и 5 % - в митохондриях, пластидах и клеточном центре в виде кольцевой ДНК.
Функции ДНК : хранение и передача информации; репарация; репликация.
Две цепи ДНК в области гена принципиально различаются по своей функциональной роли: одна из них является кодирующей, или смысловой, вторая - матричной.
Это значит, что в процессе «считывания» гена (транскрипции или синтеза пре-мРНК) в качестве матрицы выступает матричная цепь ДНК. Продукт же этого процесса-пре-мРНК - по последовательности нуклеотидов совпадает с кодирующей цепью ДНК (с заменой тиминовых оснований на урациловые).
Таким образом, получается, что с помощью матричной цепи ДНК при транскрипции воспроизводится в структуре РНК генетическая информация кодирующей цепи ДНК.
Главными матричными процессами, присущими всем живым организмам, являются репликация ДНК, транскрипция и трансляция.
Репликация - процесс, при котором информация, закодированная в последовательности оснований молекулы родительской ДНК, передается с максимальной точностью дочерней ДНК. При полуконсервативной репликации дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь является вновь синтезированной. Процесс осуществляется при участии ДНК-полимераз, которые относятся к классу трансфераз. Роль матрицы играют разделенные цепи двунитевой материнской ДНК, а субстратами являются дезоксирибонуклеозид-5"-трифосфаты.
Транскрипция - процесс переноса генетической информации от ДНК к РНК. Все виды РНК - мРНК, рРНК и тРНК - синтезируются в соответствии с последовательностью оснований в ДНК, служащей матрицей. Транскрибируется только одна, так называемая «+»-цепь ДНК. Процесс протекает при участии РНК-полимераз. Субстратами являются рибонуклеозид-5"-трифосфаты.
Процессы репликации и транскрипции у прокариот и эукариот существенно различаются по скорости протекания и по отдельным механизмам.
Трансляция - процесс декодирования мРНК, в результате которого информация с языка последовательности оснований мРНК переводится на язык аминокислотной последовательности белка. Осуществляется трансляция на рибосомах, субстратами являются аминоацил-тРНК.
Матричный синтез ДНК, катализируемый ДНК-полимеразами, выполняет две основные функции: репликацию ДНК - синтез новых дочерних цепей и репарацию двунитевых ДНК, имеющих разрывы в одной из цепей, образовавшихся в результате вырезания нуклеазами поврежденных участков этой цепи. У прокариот и эукариот существует три разновидности ДНК-полимераз. У прокариот выделены полимеразы I, II и III типов, обозначаемые как pol l, pol ll и pol III. Последняя катализирует синтез растущей цепи, pol играет важную роль в процессе созревания ДНК, функции pol ll изучены не полностью. В эукариотических клетках в репликации хромосом участвует ДНК-полимераза ά, в репарации - ДНК-полимераза β, а γ разновидность является ферментом, осуществляющим репликацию ДНК митохондрий. Эти Ферменты, независимо от типа клеток, в которых происходит репликация, присоединяют нуклеотид к ОН-группе на З"-конце одной из цепей ДНК, которая растет в направлении 5"→3. Поэтому говорят, что данные Ф обладают 5"→3"-полимеразной активностью. Помимо этого все они проявляют способность деградировать ДНК, отщепляя, нуклеотиды в направлении 3"→5, т. е. являются 3"→5"-экзонуклеазами.
В 1957 г. Мезельсон и Сталь, изучая E. coli установили, что на каждой свободной цепи фермент ДНК-полимераза строит новую, комплементарную цепь. Это полуконсервативный способ репликации: одна цепь старая – другая новая!
Обычно репликация начинается в строго определенных участках, получивших название участков ori (от origin of replication), и от этих участков распространяется в обе стороны. Участкам ori предшествуют точки разветвления материнских цепей ДНК. Участок, примыкающий к точке разветвления, получил название репликативной вилки (рис.9). В ходе синтеза репликативная вилка перемещается вдоль молекулы, при этом расплетаются все новые участки родительской ДНК до тех пор, пока вилка не дойдет до точки терминации. Разделение цепей достигается с помощью специальных Ф - геликаз (топоизомераз). Энергия, необходимая для этого, высвобождается за счет гидролиза АТФ. Геликазы перемещаются вдоль полинуклеотидных цепей в двух направлениях.
Для начала синтеза ДНК необходима затравка - праймер. Роль праймера выполняет короткая РНК (10-60 нуклеотидов). Она синтезируется комплементарно определенному участку ДНК при участии праймазы. После образования праймера в работу включается ДНК-полимераза. В отличие от геликаз ДНК-полимеразы могут перемещаться только от 3" к 5" концу матрицы. Поэтому элонгация растущей цепи по мере раскручивания двунитевой материнской ДНК может идти только вдоль одной цепи матрицы, той, относительно которой вилка репликации движется от 3" к 5" концу. Непрерывно синтезируемая цепь получила название лидирующей. Синтез на запаздывающей цепи также начинается с образования праймера и идет в направлении, противоположном ведущей цепи - от вилки репликации. Запаздывающая цепь синтезируется фрагментарно (в виде фрагментов Оказаки), т. к. праймер образуется только тогда, когда вилка репликации освободит тот участок матрицы, который имеет сродство к праймазе. Лигирование (сшивание) фрагментов Оказаки с образованием единой цепи носит название процесса созревания.
При созревании цепи РНК-затравка удаляется как с 5" конца ведущей цепи, так и с 5" концов фрагментов Оказаки, а эти фрагменты сшиваются друг с другом. Удаление затравки осуществляется при участии 3"→5" экзонуклеазы. Этот же Ф вместо удаленной РНК присоединяет дезоксинуклеотиды, используя свою 5"→3" полимеразную активность. При этом в случае присоединения «неправильного» нуклеотида осуществляется «корректорская правка» - удаление оснований, образующих некомплементарные пары. Этот процесс обеспечивает чрезвычайно высокую точность репликации, отвечающую одной ошибке на 109 пар оснований.
Рис.9. Репликация ДНК:
1 - репликативная вилка, 2 - ДНК-полимераза (pol I - созревание);
3 - ДНК-полимераза (pol III - «корректорская правка»); 4-геликаза;
5-гираза (топоизомераза); 6-белки, дестабилизирующие двойную спираль.
Коррекция осуществляется в тех случаях, когда к З"-концу растущей цепи присоединяется «неправильный» нуклеотид, неспособный образовать нужные водородные связи с матрицей. Когда pol III ошибочно присоединяет неправильное основание, «включается» ее 3" -» 5"-экзонуклеазная активность, и это основание немедленно удаляется, после чего восстанавливается полимеразная активность. Такой простой механизм действует благодаря тому, что pol III способна работать как полимераза лишь на совершенной двойной спирали ДНК с абсолютно правильным спариванием оснований.
Еще один механизм удаления РНК-фрагментов основан на присутствии в клетках особой рибонуклеазы, получившей название РНКазы Н. Этот Ф специфичен к двунитевым структурам, построенным из одной рибонуклеотидной и одной дезоксирибонуклеотидной цепи, причем он гидролизует первую из них.
РНКаза Н также способна удалять РНК-праймер с последующей застройкой разрыва с помощью ДНК-полимеразы. На заключительных этапах сборки фрагментов в нужном порядке действует ДНК-лигаза, катализирующая образование фосфодиэфирной связи.
Раскручивание геликазами части двойной спирали ДНК в хромосомах эукариот приводит к сверхспирализации остальной части структуры, что неизбежно сказывается на скорости процесса репликации. Сверхспирализации препятствуют ДНК-топоизомеразы.
Таким образом, в репликации ДНК, помимо ДНК-полимеразы, принимает участие большой набор Ф: геликаза, праймаза, РНКаза Н, ДНК-лигаза и топоизомераза. Этим перечень Ф и белков, участвующих в матричном биосинтезе ДНК, далеко не исчерпывается. Однако многие из участников этого процесса до настоящего времени остаются мало изученными.
В процессе репликации происходит «корректорская правка» - удаление неправильных (образующих некомплементарные пары) оснований, включенных во вновь синтезированную ДНК. Этот процесс обеспечивает чрезвычайно высокую точность репликации, отвечающую одной ошибке на 109 пар оснований.
Теломеры. В 1938г. классики генетики Б.Мак-Клинтон и Г. Мёллер доказали, что на концах хромосом есть специальные структуры, которые назвали теломерами (телос-конец, мерос-часть).
Ученые обнаружили, что при воздействии рентгеновским облучением устойчивость проявляют лишь теломеры. Напротив, лишенные концевых участков, хромосомы начинают сливаться, что ведет к тяжелым генетическим аномалиям. Т.о., теломеры обеспечивают индивидуальность хромосом. Теломеры плотно упакованы (гетерохроматин) и малодоступны для ферментов (теломеразы, метилазы, эндонуклеаз и др.)
Функции теломер.
1.Механические: а) соединение концов сестринских хроматид после S-фазы; б) фиксация хромосом к ядерной мембране, что обеспечивает конъюгацию гомологов.
2.Стабилизационные: а) предохранение от недорепликации генетически значимых отделов ДНК (теломеры не транскрибируются); б) стабилизация концов разорванных хромосом. У больных α - талассемией в генах α - глобина происходят разрывы хромосомы 16д и к поврежденному концу добавляются теломерные повторы (ТТАГГГ).
3.Влияние на экспрессию генов. Активность генов, расположенных рядом с теломерами, снижена. Это проявление сайленсинга – транскрипционное молчание.
4.«Счетная функция». Теломеры выступают в качестве часового устройства, которое отсчитывает количество делений клетки. Каждое деление укорачивает теломеры на 50-65 н.п. А всего их длина в клетках эмбриона человека составляет 10-15 тысяч н.п.
Теломерная ДНК попала в поле зрения биологов совсем недавно. Первые объекты исследования – одноклеточные простейшие – ресничная инфузория (тетрахимена), которая содержит несколько десятков тысяч очень мелких хромосом и, значит, множество теломер в одной клетке (у высших эукариот менее 100 теломер на клетку).
В теломерной ДНК инфузории многократно повторяются блоки из 6-ти нуклеотидных остатков. Одна цепь ДНК содержит блок 2 тимин – 4 гуанин (ТТГГГГ - Г-цепь), а комплементарная цепь - 2 аденин – 4 цитозин (ААЦЦЦЦ - Ц-цепь).
Каково же было удивление ученых, когда обнаружили, что теломерная ДНК человека отличается от таковой у инфузории всего лишь одной буквой и образует блоки 2 тимин – аденин – 3 гуанин (ТТАГГГ). Более того, оказалось, что из ТТАГГГ - блоков построены теломеры (Г – цепь) всех млекопитающих, рептилий, амфибий, птиц и рыб.
Впрочем, удивляться здесь нечему, так как в теломерной ДНК не закодировано никаких белков (она не содержит гены). У всех организмов теломеры выполняют универсальные функции, речь о которых шла выше. Очень важная характеристика теломерных ДНК – их длина. У человека она колеблется от 2 до 20 тысяч пар оснований, а у некоторых видов мышей может достигать сотен тысяч н.п. Известно, что около теломер есть специальные белки, обеспечивающие их работу и участвующие в построении теломер.
Доказано, что для нормального функционирования каждая линейная ДНК должна иметь две теломеры: по одной теломере на каждый конец.
У прокариот теломеров нет – их ДНК замкнута в кольцо.
ДНК является универсальным источником и хранителем наследственной информации, которая записана с помощью специальной последовательности нуклеотидов, она определяет свойства всех живых организмов.
Средняя молекулярная масса нуклеотида принимается равной 345, а количество нуклеотидных остатков может достигать нескольких сот, тысяч и даже миллионов. ДНК в основной своей массе находится в ядрах клеток. Немного содержится в хлоропластах и митохондриях. Однако ДНК ядра клетки - это не одна молекула. Она состоит из множества молекул, которые распределены по разным хромосомам, их количество меняется в зависимости от организма. Это и есть особенности строения ДНК.
История открытия ДНК
Строение и функции ДНК были открыты Джеймсом Уотсоном и Френсисом Криком, им даже была вручена Нобелевская премия в 1962 году.

Но впервые обнаружил нуклеиновые кислоты швейцарский ученый Фридрих Иоганн Мишер, работавший в Германии. В 1869 году он изучал животные клетки - лейкоциты. Для их получения использовал повязки с гноем, достававшиеся ему из больниц. Из гноя Мишер вымывал лейкоциты, а из них выделял белок. В ходе этих исследований ученому удалось установить, что в лейкоцитах кроме белков имеется еще что-то, какое-то неизвестное на тот момент вещество. Оно представляло собой нитевидный или хлопьевидный осадок, который выделялся, если создать кислую среду. Осадок сразу растворялся при добавлении щелочи.

Ученый с помощью микроскопа обнаружил, что при отмывании лейкоцитов с помощью соляной кислоты от клеток остаются ядра. Тогда он сделал заключение, что в ядре есть неизвестное вещество, названное им нуклеином (слово nucleus в переводе означает ядро).
Проведя химический анализ, Мишер выяснил, что новое вещество в своем составе имеет углерод, водород, кислород и фосфор. В то время фосфорорганических соединений было известно немного, поэтому Фридрих решил, что обнаружил новый класс соединений, находящихся в ядре клетки.
Таким образом, в XIX веке было открыто существование нуклеиновых кислот. Однако в то время никто не мог даже подумать о том, какая важная роль им принадлежит.
Вещество наследственности
Строение ДНК продолжали исследовать, и в 1944 году группа бактериологов под руководством Освальда Эвери получила доказательства того, что эта молекула заслуживает серьезного внимания. Ученый на протяжении многих лет занимался изучением пневмококков, организмов, которые вызывали пневмонию или заболевание легких. Эвери проводил опыты, смешивая пневмококки, вызывающие заболевание, с теми, которые безопасны для живых организмов. Сначала болезнетворные клетки убивали, а после добавляли к ним те, которые заболеваний не вызывают.
Результаты исследований поразили всех. Были такие живые клетки, которые после взаимодействия с мертвыми научались вызывать болезнь. Ученый выяснил природу вещества, которое участвует в процессе передачи информации живым клеткам от мертвых. Молекула ДНК и оказалась этим веществом.

Строение
Итак, необходимо разобраться с тем, какое строение имеет молекула ДНК. Открытие ее структуры стало значимым событием, это привело к образованию молекулярной биологии - новой отрасли биохимии. ДНК в больших количествах находится в ядрах клеток, однако размеры и количество молекул зависят от вида организма. Установлено, что ядра клеток млекопитающих содержат много этих клеток, они распределены по хромосомам, их насчитывается 46.
Изучая строение ДНК, в 1924 году Фельген впервые установил ее локализацию. Доказательства, полученные в ходе экспериментов, показали, что ДНК находится в митохондриях (1-2%). В других местах эти молекулы могут находиться при вирусной инфекции, в базальных тельцах, а также в яйцеклетках некоторых животных. Известно, что чем сложнее организм, тем масса ДНК больше. Количество молекул, находящихся в клетке, зависит от функции и составляет обычно 1-10%. Меньше всего их находится в миоцитах (0,2%), больше - в половых клетках (60%).

Строение ДНК показало, что в хромосомах высших организмов они связаны с простыми белками - альбуминами, гистонами и прочими, которые все вместе образуют ДНП (дезоксирибонуклеопротеид). Обычно большая молекула нестойкая, и для того чтобы она оставалась целой и неизменной в ходе эволюции, создана так называемая репарирующая система, которая состоит из ферментов - лигаз и нуклеаз, отвечающих за «ремонт» молекулы.
Химическое строение ДНК
ДНК является полимером, полинуклеотидом, состоящим из огромного числа (до десятков тысяч миллионов) мононуклеотидов. Строение ДНК имеет следующий вид: мононуклеотиды содержат азотистые основания - цитозин (Ц) и тимин (Т) - из производных пиримидинов, аденин (А) и гуанин (Г) - из производных пурина. Кроме азотистых оснований, в составе молекулы человека и животных имеется 5-метилцитозин — минорное пиримидиновое основание. С фосфорной кислотой и дезоксирибозой связываются азотистые основания. Схема строения ДНК продемонстрирована ниже.

Правила Чаргаффа
Строение и биологическая роль ДНК изучались Э. Чаргаффом в 1949 году. В ходе исследований он выявил закономерности, которые наблюдаются в количественном распределении азотистых оснований:
- ∑Т + Ц = ∑А + Г (то есть число пиримидиновых оснований равно числу пуриновых).
- Всегда количество остатков аденина равно количеству остатков тимина, а количество гуанина равно цитозину.
- Коэффициент специфичности имеет формулу: Г+Ц/А+Т. Например, у человека он равен 1,5, у быка - 1,3.
- Сумма "А + Ц" равна сумме "Г + Т", то есть аденина и цитозина имеется столько же, сколько гуанина и тимина.
Модель строения ДНК
Ее создали Уотсон и Крик. Остатки фосфатов и дезоксирибоз располагаются по хребту двух закрученных спиралеобразным образом полинуклеотидных цепей. Определено, что плоскостные структуры пиримидиновых и пуриновых оснований располагаются перпендикулярно оси цепи и образуют как бы ступени лестницы в виде спирали. Установлено также, что А всегда соединяется с Т при помощи двух водородных связей, а Г прикреплено к Ц уже тремя такими же связями. Этому явлению дали название "принцип избирательности и комплементарности".
Уровни структурной организации
Изогнутая как спираль полинуклеотидная цепь - это первичная структура, которая имеет определенный качественный и количественный набор мононуклеотидов, связанных 3’,5’-фосфодиэфирной связью. Таким образом, каждая из цепей имеет 3’-конец (дезоксирибоза) и 5’-конец (фосфатный). Участки, которые содержат в себе генетическую информацию, названы структурными генами.
Двухспиральная молекула - это вторичная структура. Причем ее полинуклеотидные цепи антипараллельны и связываются водородными связями между комплементарными основаниями цепей. Установлено, что в каждом витке этой спирали содержится 10 нуклеотидных остатков, длина ее равняется 3,4 нм. Эту структуру поддерживают также Ван-дер-Ваальсовы силы взаимодействия, которые наблюдаются между основаниями одной цепи, включающие отталкивающие и притягивающие компоненты. Эти силы объясняются взаимодействием электронов в соседних атомах. Электростатическое взаимодействие также стабилизирует вторичную структуру. Оно возникает между заряженными положительно молекулами гистонов и заряженной отрицательно нитью ДНК.
Третичная структура - это намотка цепей ДНК на гистоны или суперспирализация. Описано пять видов гистонов: Н1, Н2А, Н2В, Н3, Н4.
Укладка нуклеосом в хроматин - это четвертичная структура, поэтому молекула ДНК, имеющая длину несколько сантиметров, может складываться до 5 нм.

Функции ДНК
Основными функциями ДНК являются:
- Хранение наследственной информации. Последовательность аминокислот, находящихся в молекуле белка, определяется порядком, в котором расположены нуклеотидные остатки в молекуле ДНК. Также в ней зашифрована вся информация о свойствах и признаках организма.
- ДНК способна передавать наследственную информацию следующему поколению. Это возможно из-за способности к репликации - самоудвоению. ДНК способна распадаться на две комплементарные цепочки, и на каждой из них (в соответствии с принципом комплементарности) восстанавливается исходная последовательность нуклеотидов.
- При помощи ДНК происходит биосинтез белков, ферментов и гормонов.
Заключение
Строение ДНК позволяет ей являться хранителем генетической информации, а также передавать ее следующим поколениям. Какие есть особенности у этой молекулы?
- Стабильность. Это возможно благодаря гликозидным, водородным и фосфодиэфирным связям, а также механизму репарации индуцированных и спонтанных повреждений.
- Возможность репликации. Этот механизм позволяет в соматических клетках сохранять диплоидное число хромосом.
- Существование генетического кода. При помощи процессов трансляции и транскрипции последовательность оснований, находящихся в ДНК, преобразуется в последовательность аминокислот, находящихся в полипептидной цепи.
- Способность к генетической рекомбинации. При этом образуются новые сочетания генов, которые сцеплены между собой.
Таким образом, строение и функции ДНК позволяют ей играть неоценимую роль в организмах живых существ. Известно, что длина 46-ти молекул ДНК, находящихся в каждой клетке человека, равна почти 2 м, а число нуклеотидных пар составляет 3,2 млрд.
Свойства ДНК определяются ее строением:
1. Универсальность - принципы построения ДНК для всех организмов одинаковы.
2. Специфичность - определяется соотношением азотистых оснований: А + Т,
которое специфично для каждого вида. Так у человека оно составляет 1,35, у бактерий – 0,39
Специфичность зависит от:
· количества нуклеотидов
· вида нуклеотидов
· расположение нуклеотидов в цепи ДНК
2. Репликация или самоудвоение ДНК: ДНК↔ДНК. Генетическая программа клеточных организмов записана в нуклеотидной последовательности ДНК. Для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Во время деления клетки содержание ДНК должно удвоиться, чтобы каждая дочерняя клетка могла получить полный спектр ДНК, т.е. в любой делящейся соматической клетке человека должно быть скопировано 6,4*10 9 нуклеотидных пар. Процесс удвоения ДНК получил название репликации. Репликация относится к реакциям матричного синтеза. Во время репликации каждая из двух цепей ДНК служит матрицей для образования комплементарной (дочерней) цепи. Протекает она в S-период интерфазы клеточного цикла. Высокая надежность процесса репликации гарантирует практически безошибочную передачу генетической информации в ряду поколений. Пусковым сигналом для начала синтеза ДНК в S-периоде является так называемый S – фактор (специфические белки). Зная скорость репликации и длину хромосомы эукариот можно рассчитать время репликации, которое теоретически составляет несколько суток, а практически репликация осуществляется за 6 – 12 часов. Из этого следует, что репликация у эукариот одновременно начинается в нескольких местах на одной молекуле ДНК.
Единицей репликации является репликон. Репликон – это участок ДНК, где происходит репликация. Количество репликонов на одну интерфазную хромосому у эукариот может достигать 100 и более. В клетке млекопитающих может быть 20 – 30 тыс. репликонов, у человека – примерно 50 тыс. При фиксированной скорости роста цепи (у эукариот – 100 нуклеотидов в секунду) множественная инициация обеспечивает большую скорость процесса и снижение времени, необходимого для дупликации протяженных участков хромосом, т.е. у эукариот осуществляется полирепликонная репликация. (рис. 21)
Репликон содержит все необходимые гены и регуляторные последовательности, которые обеспечивают репликацию. Каждый репликон в процессе клеточного деления активируется один раз. Репликация контролируется на стадии инициации. Если процесс удвоения начался он будет продолжаться до тех пор, пока весь репликон не будет удвоен.
У прокариот вся ДНК является одним репликоном.
Рис.21. Репликация хромосомной ДНК эукариот. Репликация идет в двух направлениях из разных точек начала репликации (Ori) с образованием пузырьков. «Пузырь» или «глаз» это область реплицированной ДНК внутри нереплицированной. (А. С. Коничев, Г. А. Севастьянова, 2005, с. 213)
Ферменты, участвующие в процессе репликации, объединены в мультиферментативный комплекс . В репликации ДНК у прокариот участвует 15 ферментов, а у эукариот – более 30, т.е. репликация – это архисложный и суперточный многоступенчатый ферментативный процесс. В состав ферментативных комплексов входят следующие ферменты:
1) ДНК – полимеразы (I, III), катализируют комплементарное копирование, т.е. отвечают за рост дочерней цепи. (рис. 22) Прокариоты реплицируются со скоростью 1000 нуклеотидов в секунду, а эукариоты - 100 нуклеотидов в секунду. Пониженная скорость синтеза у эукариот связана с затрудненной диссоциацией гистоновых белков, которые необходимо удалить для продвижения ДНК-полимеразы в репликативной вилке вдоль цепи ДНК.
2) ДНК - праймаза. ДНК – полимеразы могут удлинять полинуклеотидную цепь присоединяясь к уже имеющимся нуклеотидам. Поэтому, чтобы ДНК – полимераза смогла начать синтез ДНК, ей необходима затравка или праймер (от. англ. primer – затравка). ДНК – праймаза синтезирует такую затравку, которая затем замещается сегментами ДНК. (рис. 22).
3) ДНК – лигаза, соединяет фрагменты Оказаки друг с другом за счет образования фосфодиэфирной связи.
4) ДНК – хеликаза, расплетает спираль ДНК, разрывает водородные связи между ними. В результате образуются две одиночные разнонаправленные ветви ДНК (рис.22).
5) SSB – белки, связываются с одноцепочечной ДНК и стабилизируют её, т.е. они создают условия для комплементарного спаривания.
Репликация ДНК начинается не в любой случайной точке молекулы, а в специфических местах, называемых областью (точками) начала репликации (Ori). Они имеют определенные последовательности нуклеотидов, что облегчает разделение цепей (рис.21). В результате инициации репликации в точке Ori образуются одна или две репликативные вилки – места разделения материнских цепей ДНК. Процесс копирования продолжается до тех пор, пока ДНК полностью не удвоится или пока репликативные вилки двух соседних точек начала репликации не сольются. Точки начала репликации у эукариот разбросаны по хромосоме на расстоянии равном 20 000 пар нуклеотидов (рис.21).
Рис.22. Репликация ДНК (объяснение в тексте). (Б. Альбертс и др., 1994, т. 2, с. 82)
Фермент – хеликаза – разрывает водородные связи, т.е. расплетает двойную цепь, образуя две разнонаправленные ветви ДНК (рис.22). Одноцепочечные участки связываются специальными SSВ-белками , которые выстраиваются снаружи каждой материнской цепи и оттягивают их друг от друга. Это делает азотистые основания доступными для связывания с комплементарными нуклеотидами. В месте схождения этих ветвей по направлению репликации ДНК располагается фермент ДНК-полимераза, который катализирует процесс и контролирует точность комплементарного синтеза. Особенностью работы данного фермента является его однонаправленность, т.е. построение дочерней цепи ДНК идет по направлению от 5" конца к 3" . На одной материнской цепи синтез дочерней ДНК идет непрерывно (лидирующая цепь). Она растет от 5" к 3" концу в направлении движения репликативной вилки и поэтому нуждается только в одном акте инициации. На другой материнской цепи синтез дочерней цепи идет в виде коротких фрагментов с обычной 5" - 3" полярностью и при помощи ферментов – лигаз происходит их сшивание в одну неперывную отстающую цепь. Поэтому для синтеза отстающей цепи требуется несколько актов (точек) инициации.
Такой способ синтеза назван прерывистой репликацией. Фрагментные участки, синтезированные на отстающей цепи, в честь первооткрывателя названы фрагментами Оказаки . Они обнаружены у всех реплицирующихся ДНК, как у прокариот, так и у эукариот. Их длина соответствует 1000 – 2000 нуклеотидам у прокариот и 100 – 200 у эукариот. Таким образом, в результате репликации образуются 2 идентичные молекулы ДНК, в которых одна цепь материнская, другая вновь синтезированная. Такой способ репликации называют полуконсервативным. Предположение о таком способе репликации было сделано Дж. Уотсоном и Ф. Криком, а доказано в 1958г. М . Мезелсоном и Ф. Сталем . После репликации хроматин представляет собой систему из 2 декомпактизированных молекул ДНК, объединенных центромерой.
В процессе репликации могут возникать ошибки, которые у прокариот и эукариот бывают с одной и той же частотой - одна на 10 8 -10 10 нуклеотидов , т.е. в среднем 3 ошибки на геном . Это доказательство высокой точности и скоординированности процессов репликации.
Ошибки репликации исправляются ДНК-полимеразой III («механизм корректорской правки») или системой репараций.
2. Репарация - это свойство ДНК восстанавливать свою целостность, т.е. исправлять повреждения. Передача наследственной информации в неискаженном виде важнейшее условие выживания как отдельного организма, так и вида в целом. Большинство изменений вредны для клетки, они либо приводят к мутациям, либо блокируют репликацию ДНК, либо вызывают гибель клетки. ДНК постоянно подвергается действию спонтанных (ошибки репликации, нарушение структуры нуклеотида и т.д.) и индуцированных (УФ – облучение, ионизирующая радиация, химические и биологические мутагены) факторов среды. В ходе эволюции выработалась система позволяющая исправлять нарушения в ДНК – система репарации ДНК . В результате её активности на 1000 повреждений ДНК только одно приводит к мутациям. Повреждение - любое изменение ДНК, которое вызывает отклонение от обычной двуцепочечной структуры:
1) появление одноцепочечных разрывов;
2) удаление одного из оснований, в результате чего его гомолог остается неспаренным;
3) замещение одного основания в комплементарной паре другим, неправильно спаренным с основанием-партнером;
4) появление ковалентных связей между основаниями одной цепи ДНК или между основаниями на противоположных цепях.
Репарация может проходить до удвоения ДНК (дорепликативная репарация) и после удвоения ДНК (пострепликативная). В зависимости от характера мутагенов и степени повреждения ДНК в клетке идет световая (фотореактивация), темновая, SOS-репарация и др.
Считают, что фотореактивация идет в клетке, если повреждения ДНК вызваны естественными условиями (физиологические особенности организма, обычные факторы среды, в том числе - ультрафиолетовые лучи). Восстановление целостности ДНК при этом, происходит с участием видимого света: репаративный фермент активируется квантами видимого света, соединяется с поврежденной ДНК, разъединяет пиримидиновые димеры нарушенного участка и восстанавливает целостность нити ДНК.
Темновая репарация (эксцизионная) наблюдается после действия ионизирующей радиации, химических веществ и т.д. Она включает удаление поврежденного участка, восстановление нормальной структуры молекулы ДНК (рис.23). Для этого типа репарации необходима вторая комплементарная цепь ДНК. Темновая репарация многоступенчата, в ней участвует комплекс ферментов, а именно:
1)фермент, узнающий поврежденный участок цепи ДНК
2)ДНК – эндонуклеаза, делает разрыв в поврежденной цепи ДНК
3) экзонуклеаза удаляет измененную часть нити ДНК
4) ДНК – полимераза I синтезирует новый участок ДНК взамен удаленного
5)ДНК- лигаза сшивает конец старой нити ДНК с вновь синтезированной, т.е. замыкает два конца ДНК (рис.23). В темновой репарации у человека принимают участие 25 белков-ферментов.
При больших повреждениях ДНК, которые угрожают жизни клеток, включается SOS-репарация . SOS-репарация была открыта в 1974 году. Такой тип репарации отмечают после действия больших доз ионизирующей радиации. Характерная черта SOS-репарации - неточность восстановления первичной структуры ДНК, в связи с чем она получила название репарации, склонной к ошибкам . Главная цель SOS-репарации сохранить жизнеспособность клетки.
Нарушение в системе репарации могут приводить к преждевременному старению, развитию онкологических заболеваний, болезням аутоиммунной системы, гибели клетки или организма.
Рис. 23. Репарация поврежденной ДНК путем замены модифицированных нуклеотидных остатков (темновая репарация или эксцизионная). (М. Сингер, П. Берг, 1998, т. 1, с.100)